Многочисленные опыты подтвердили правильность установленных Менделем закономерностей. Вместе с тем, появились факты, показывающие, что полученные Менделем числовые соотношения при расщеплении гибридного поколения соблюдались не всегда. Это указывало на то, что взаимоотношения между генами и признаками носят более сложный характер. Выяснилось: один и тот же ген может оказывать влияние на развитие нескольких признаков; один и тот же признак может развиваться под влиянием многих генов .
Следует отметить, что взаимодействие генов имеет биохимическую природу, то есть взаимодействуют друг с другом не гены, а их продукты. Продуктом эукариотического гена может быть или полипептид, или тРНК, или рРНК.
ВИДЫ ВЗАИМОДЕЙСТВИЯ АЛЛЕЛЬНЫХ ГЕНОВ
Различают полное доминирование, неполное доминирование, кодоминирование, аллельное исключение.
Аллельными генами называются гены, расположенные в идентичных локусах гомологичных хромосом. Ген может иметь одну, две и более молекулярных форм. Появление второй и последующих молекулярных форм является следствием мутации гена. Если ген имеет три и более молекулярных форм, говорят о множественном аллелизме . Из всего множества молекулярных форм у одного организма могут присутствовать только две, что объясняется парностью хромосом.
Полное доминирование
Полное доминирование — это вид взаимодействия аллельных генов, при котором фенотип гетерозигот не отличается от фенотипа гомозигот по доминанте, то есть в фенотипе гетерозигот присутствует продукт доминантного гена. Полное доминирование широко распространено в природе, имеет место при наследовании, например, окраски и формы семян гороха, цвета глаз и цвета волос у человека, резус-антигена и мн. др.
Наличие резус-антигена (резус-фактора) эритроцитов обусловливается доминантным геном Rh . То есть генотип резус-положительного человека может быть двух видов: или RhRh , или Rhrh ; генотип резус-отрицательного человека — rhrh . Если, например, мать — резус-отрицательная, а отец резус-положительный и гетерозиготен по этому признаку, то при данном типе брака с одинаковой вероятностью может родиться как резус-положительный, так и резус-отрицательный ребенок.
Между резус-положительным плодом и резус-отрицательной матерью может возникнуть резус-конфликт.

Так называется вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву и имеет среднее (промежуточное) значение между ними. Имеет место при наследовании окраски околоцветника ночной красавицы, львиного зева, окраски шерсти морских свинок и пр.
Сам Мендель столкнулся с неполным доминированием, когда скрещивал крупнолистный сорт гороха с мелколистным. Гибриды первого поколения не повторяли признак ни одного из родительских растений, они имели листья средней величины.
При скрещивании гомозиготных красноплодных и белоплодных сортов земляники все первое поколение гибридов имеет розовые плоды. При скрещивании этих гибридов друг с другом получаем: по фенотипу — 1/4 красноплодных, 2/4 розовоплодных и 1/4 белоплодных растений, по генотипу — 1/4 АА , 1/2 Аа , 1/4 аа (и по фенотипу, и по генотипу соотношение 1:2:1). Соответствие расщепления по генотипу расщеплению по фенотипу является характерным при неполном доминировании, так как гетерозиготы фенотипически отличаются от гомозигот.
Кодоминирование
Кодоминирование — вид взаимодействия аллельных генов, при котором фенотип гетерозигот отличается как от фенотипа гомозигот по доминанте, так и от фенотипа гомозигот по рецессиву, и в фенотипе гетерозигот присутствуют продукты обоих генов. Имеет место при формировании, например, IV группы крови системы (АВ0) у человека.
Для того чтобы представить, как происходит наследование групп крови у человека, можно посмотреть, рождение детей с какой группой крови возможно у родителей, имеющих один — вторую, другой — третью группы крови и являющихся гетерозиготными по этому признаку.
| Р | ♀I A i 0
II (A) |
× | ♂I B i 0
III (B) |
||
| Типы гамет | I A | i 0 | I B | i 0 | |
| F | i 0 i 0
I (0) 25% |
I A i 0
II (A) 25% |
I B i 0
III (B) 25% |
I A I B
IV (AB) 25% |
|
Аллельное исключение
Аллельным исключением называется отсутствие или инактивация одного из пары генов; в этом случае в фенотипе присутствует продукт другого гена (гемизиготность, делеция, гетерохроматизация участка хромосомы, в котором находится нужный ген).
ВИДЫ ВЗАИМОДЕЙСТВИЯ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Комплементарность, эпистаз, полимерия.
Неаллельные гены — гены, расположенные или в неидентичных локусах гомологичных хромосом, или в разных парах гомологичных хромосом.
Наблюдение между аллельными генами; гетерозиготы проявляют признаки промежуточные по отношению к родительским. При неполном доминирование в потомстве гибридов (F2) расщепление по генотипу и фенотипу совпадает (1:2:1)
Пример:окраска цветков ночной красавицы.
Генетическое заболевание человека – брахидактилия - укорочение средней фаланги пальцев. За развитие заболевания отвечает ген «в», здоровый человек имеет генотип «ВВ». Больные брахидактилией являются гетерозиготами и имеют генатип «Вв». Гомозиготы по гену «вв» погибают до рождения, то есть ген «в» в гомозиготном состояние летален.
Кодоминантность – независимое проявление аллельных генов. Каждый в генотипе проявляет свое действие, а в результате проявляют новый признак.
Пример: формирование четвертой группы крови у человека. Наследование группы крови у человека по системе АВО, обусловлено геном I, существует 3 аллели этого гена. I О; I А; I В. Гены I A и I B доминантны по отношению к I O , но кодоминантны по отношению друг к другу, и у лиц имеющих генотип I A I B проявляется 4 группа крови.
Комплементарност ь – проявление признака происходящего лишь в случае присутствия в генотипе организма двух доминантных неаллельных генов.
Пример: комплементарное взаимодействия наблюдений при наследовании форм гребня у кур.
 Эпистаз
– взаимодействие неаллельных генов, при котором ген одной аллельной пары подавляет действие другой аллельной пары. Ген, подавляющий действие другого гена называется эпистатическим
, супрессором или ингибирующим. Эпистаз может быть доминантным и рецессивным.
Эпистаз
– взаимодействие неаллельных генов, при котором ген одной аллельной пары подавляет действие другой аллельной пары. Ген, подавляющий действие другого гена называется эпистатическим
, супрессором или ингибирующим. Эпистаз может быть доминантным и рецессивным.
Пример: доминантные эпистаз: наследование окраски определения у кур. Доминантный ген С – нормальная продукция пигмента; с – не обеспечивает синтез фермента; доминантный ген I другой аллельной пары – супрессор. В результате куры имеющие в генотипе С и I, оказываются белыми.
Пример: рецессивного эпистаза служит окраска шерсти у домовых мышей
Полимерия – явление одновременного действия на признак нескольких неаллельных однотипных генов.
Пример: наследование количественных признаков: окраска семян пшеницы, кожи человека и др; яйценостность, содержание сахара в сахарной свекле и др.
Гены отвечают за развитие количественных признаков, обладают суммарным эффектом, чем больше доминированных генов, тем ярче признак.
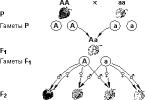
Пример: за пигментацию кожи человека отвечает полимерные гены S 1 S 1 S 2 S 2 чем больше доминирующих генов в генотипе, тем темнее кожа.
Темная кожа Светлая кожа
Р: S 1 S 1 S 2 S 2 х s 1 s 1 s 2 s 2
F1: S 1 s 1 S 2 s 2
От брака между мулатами родятся дети с пигментацией кожи от светло до темной. Вероятность рождения ребенка с черным цветом кожи равна 1/16.
ИЗМЕНЧИВОСТЬ.
Изменчивость – способность дочерних организмов отличаться от родительских форм.
Изменчивость
Ненаследственная Наследственная
(фенотипическая, (генотипическая,
модификационная, индивидуальная,
направленная) случайная)
Комбинативная Мутационная
Модификационная изменчивость (ненаследственная) – представляет собой эволюционно закреплённые приспособленческие реакции организма на изменения условий внешней среды без изменения генотипа.
Характеристика – затрагивает только фенотип; не передается по наследству; определяется условиями существования; носит приспособительный характер к условиям среды.
Модификация (от лат. «modificatio» - «видоизменение») – ненаследственное изменение фенотипа, возникающее под влиянием факторов внешней среды в пределах нормы реакции генотипа.
Пределы, в которых возможно изменение признаков у данного генотипа, называется нормой реакции. По степени варьирования нормы реакции признаки бывают пластичные и непластичные. Пластичные (широкая норма реакции), примеры: жирность молока у коров, масса тела человека, и т. д. Непластичные (узкая норма реакции), примеры – окраска радужки глаз, группа крови у человека, и т.п.
Примеры модификационной изменчивости:
Количество эритроцитов в 1 мл 3 крови увеличивается почти вдвое у альпинистов, поднимающихся на высоту 4000 м.
Сосна, выросшая на горном склоне, будет сильно внешне отличаться по росту и форме ствола от такой же сосны, растущей на равнине.
У людей индивидуальность и талант проявляются в результате воздействия генетических задатков и внешней среды, причём решающая роль в этом принадлежит воспитанию и общественным отношениям.
Модификационная изменчивость характеризуется следующими особенностями:
ü групповой характер изменений:например, находясь под воздействием УФ-лучей, все отдыхающие на пляже загорают, но интенсивность загара отличается.
ü адекватность изменений:чем больше люди находились под воздействием лучей солнца, тем больше загорали.
ü кратковременность большинства модификаций: люди приезжают с юга, и через несколько недель загар сходит.
ü адаптивность изменений:загар – защита организма от действия УФ-лучей.
ü ограниченность: - норма реакции.
Признаки организма могут быть:
Качественными (цвет глаз и волос у человека);
Количественными (рост и масса тела у человека).
Для характеристики степени изменчивости количественных признаков применяется один из методов статистики – построение вариационной кривой.
Дарвин называл модификацию изменений определённой, т.к. все особи одного вида, попав в сходные условия, изменяются одинаково, т.е., такая изменчивость предсказуема, например: все овцы при выращивании в более холодных условиях стали иметь более густую шерсть.
Генотипической называется изменчивость самого генотипа организма.
Характеристика: передаётся по наследству; затрагивает генотип; носит случайный характер.Подразделяется на комбинативную и мутационную.
Комбинативная изменчивость связана с получением новых комбинаций имеющихся в генотипе генов. Обусловлена: независимым расхождением хромосом в мейозе; случайным сочетанием хромосом при оплодотворении; перекомбинацией генов при кроссинговере.
Пример: появление зелёной окраски семян гороха при скрещивании гетерозиготных растений с растениями с жёлтыми семенами.
Мутационная изменчивость – изменение самого генотипа, в результате мутаций.
Мутации – внезапные скачкообразные и ненаправленные изменения ДНК, с появлением у живых организмов качественно новых признаков и свойств, которых ранее в природе не существовало.
Основные положения мутационной теории разработаны Г. Де Фризоне.
Мутации возникают внезапно, как дискретные изменения признаков;
Новые фенотипы устойчивы;
В отличие от ненаследственных изменений, мутации не образуют непрерывных рядов, представляющих собой качественные изменения;
Мутации могут быть вредными, нейтральными и полезными для организма;
Вероятность обнаружения мутаций зависит от числа исследуемых особей;
Сходные мутации могут возникать повторно.
Причины мутаций – воздействие мутационных факторов различного происхождения. Их подразделяют на:
ü Физические (ионизирующая радиация: альфа-, бета-, гамма-излучения, УФ-лучи, высокая температура);
ü Химические (формалин, иприт, лекарства, пищевые консерванты, пестициды и др.);
ü Биологические (вирусы, бактерии).
Генные мутации связаны с изменением нуклеотидной последовательности ДНК одного гена. Типы генных мутаций проявляются в формах: дупликаций (повторение набора генов, локализованных в этом участке), вставок, делеций (выпадение участков хромосом в средней части), инверсий (поворот участка на 180 о), дефименси (потеря концевых участков хромосом), транслокации (перенос участка к другому концу той же хромосомы либо к другой негомологичной хромосоме).
Пример: дефименси – синдром «кошачьего крика» - гетерозиготность по дефименси в пятой хромосоме.
Хромосомные мутации связаны с перемещеним участков хромосом.
Геномны е мутации – изменение числа хромосом в геноме клетки (в кариотипе особи).
Геном – содержание наследственного материала в гаплоидном наборе хромосом.
К геномным мутациям относятся:
Полиплоидия (эуплоидия) – кратное увеличение гаплоидного набора хромосом.
Клетки с различным числом гаплоидных наборов называются триплоидными (3), тетраплоидными (4), гексаплоидными (6), и т.п.
Полиплоиды образуются при нарушении расхождения хромосом к полюсам клетки в митозе. Полиплоидия распространена главным образом у растений. Полиплоидные формы имеют более крупные листья, цветы, плоды и семена. Многие культурные растения являются полиплоидами. Существует 2 типа полиплоидии: аутополиплоидия и аллополиплоидия.
Гетероплоидия (анэуплоидия) – вид геномной мутации, при которой происходит некратное гаплоидному увеличение или изменение количества хромосом. (2n-1 – моносомия, 2n+1 – трисомия; полисомия и др.).
У человека анэуплодия приводит к бесплодию и часто к хромосомным болезням (синдром Дауна 2n = 47; синдром Шериневского-Тернера, синдром Клайнфельтера и др.).
Мутации классифицируются:
1) по причинам, вызвавшим мутацию: спонтанные (в естественных условиях) и индуцированные (под направленным воздействием на органы мутационных факторов). Впервые данные мутации были получены Г. А. Надсоном и Л. С. Филипповым (1925г.) при облучении грибов радием, и Г. Меллером (1927г.) при облучении мух-дрозофил рентгеновскими лучами.
2) по характеру мутировавших клеток: соматические – проявляются у самой особи, не наследуются при половом размножении, наследуются при вегетативном. Пример: разный цвет радужки глаз у человека) и генеративные – происходят в половых клетках, наследуются, выявляются фенотипически у потомков, являющихся материалом для естественного отбора.
3) по исходу для организма: отрицательные – летальные/полулетальные (снижение жизнеспособности); нейтральные; положительные (встречаются редко).
СЕЛЕКЦИЯ
Селекция (от лат. “selection” – «отбор», «выбор») – это получение новых сортов растений, пород животных и штаммов микроорганизмов с ценными для человека свойствами.
Породой, сортом, штаммом называют популяцию организмов, искусственно созданную человеком и характеризующуюся определёнными наследственными особенностями.
Теоретической основой селекции является генетика.
Основными методами селекции являются:
гибридизация;
полиплоидия;
мутагенез;
клеточная и генная инженерия.
Н. И. Вавилов – сформулировал закон гомологических рядов в наследственной изменчивости; учение о нахождении материала для селекции является созданное им представление о центрах происхождения культурных растений. Он определил 7 таких центров.
Значительный вклад в селекцию плодовых культур внёс И. В. Мичурин. Он исполнил методы гибридизации, отбора и воздействия условиями среды («метод ментора») на развивающиеся гибриды. Важное место в селекционных работах Мичурина занимало управление доминированием, которое основано на представлении о том, что в конкретных условиях среды у гибридов преимущественное развитие получают благоприятные для данных условий признаки
При неполном доминировании гетерозиготы имеют фенотип, промежуточный между фенотипами доминантной и рецессивной гомозиготы. Например, при скрещивании чистых линий львиного зева и многих других видов цветковых растений с пурпурными и белыми цветками особи первого поколения имеют розовые цветки. При скрещивании чистых линий андалузских кур чёрной и белой окраски в первом поколении рождаются куры серой окраски. На молекулярном уровне самым простым объяснением неполного доминирования может быть как раз двукратное снижение активности фермента или другого белка (если доминантный аллель дает функциональный белок, а рецессивный - дефектный). Например, за белую окраску может отвечать дефектный аллель, который дает неактивный фермент, а за красную - нормальный аллель, который дает фермент, производящий красный пигмент. При половинной активности этого фермента у гетерозигот количество красного пигмента снижается вдвое, и окраска розовая. Могут существовать и другие механизмы неполного доминирования.
Иногда расщепление признаков во втором поколении может отклоняться от ожидаемых (3:1 - при полном доминировании, 1:2:1 - при неполном доминировании) результатов. Это связано с тем, что в некоторых случаях гомозиготы по одному из признаков оказываются нежизнеспособными. В этом случае говорят о летальных генах. Один ген может оказывать влияние на другие признаки, в результате чего снижается работоспособность. Например, серые каракульские овцы, гомозиготные по доминантному признаку серой окраски, погибают после рождения из-за недоразвития желудка. Другим примером доминантного летального гена является брахидактилия у человека (укороченные пальцы). Гомозиготы по данному гену погибают на ранних стадиях развития зародыша, а признак проявляется только у гетерозигот.
Примером рецессивного летального гена является ген серповидно - клеточной анемии у человека. В норме эритроциты человека имеют форму двояковогнутого диска. При серповидно - клеточной анемии они приобретают вид серпа, а физиологический эффект выражается острой анемией и снижением количества кислорода, переносимого кровью. У гетерозигот заболевание не проявляется, эритроциты тем не менее все же имеют измененную форму. Гомозиготы по этому признаку в 95% случаев гибнут в раннем возрасте из - за кислородной недостаточности, а гетерозиготы вполне жизнеспособны.
При скрещивании между собой особей F1 отношение фенотипов в F2 отличается от менделевского отношения 3: 1, типичного для моногибридного скрещивания. В этом случае получается отношение 1: 2: 1, где у половины особей F2 будет такой же генотип, как у F1. Отношение 1: 2: 1 характерно для результатов скрещиваний при неполном доминировании.
32. Генетика пола. Сцепленное с полом наследование
Генетика пола.
Пол - совокупность признаков, по которым производится специфическое разделение особей или клеток, основанное на морфологических и физиологических особенностях, позволяющее осуществлять в процессе полового размножения комбинирование в потомках наследственных задатков родителей.
Морфологические и физиологические признаки, по которым производится специфическое разделение особей, называется половым.
Признаки, связанные с формированием и функционированием половых клеток, называется первичными половыми признаками. Это гонады (яичники или семенники), их выводные протоки, добавочные железы полового аппарата, копулятивные органы. Все другие признаки, по которым один пол отличается от другого, получили название вторичных половых признаков. К ним относят: характер волосяного покрова, наличие и развитие молочных желез, строение скелета, тип развития подкожной жировой клетчатки, строение трубчатых костей и др.
Наследование, сцепленное с полом - наследование какого-либо гена, находящегося в половых хромосомах. Наследование признаков, проявляющихся только у особей одного пола, но не определяемых генами, находящимися в половых хромосомах, называется наследованием, ограниченным полом.
Наследованием, сцепленным с X-хромосомой, называют наследование генов в случае, когда мужской пол гетерогаметен и характеризуется наличием Y-хромосомы (XY), а особи женского пола гомогаметны и имеют две X-хромосомы (XX). Таким типом наследования обладают все млекопитающие (в том числе человек), большинство насекомых и пресмыкающихся.
Наследованием, сцепленным с Z-хромосомой, называют наследование генов в случае, когда женский пол гетерогаметен и характеризуется наличием Z-хромосомы (ZW), а особи мужского пола гомогаметны и имеют две Z-хромосомы (ZZ). Таким типом наследования обладают все представители класса птиц.
Если аллель сцепленного с полом гена, находящегося в X-хромосоме или Z-хромосоме, является рецессивным, то признак, определяемый этим геном, проявляется у всех особей гетерогаметного пола, которые получили этот аллель вместе с половой хромосомой, и у гомозиготных по этому аллелю особей гомогаметного пола. Это объясняется тем, что вторая половая хромосома (Y или W) у гетерогаметного пола не несет аллелей большинства или всех генов, находящихся в парной хромосоме.
Таким признаком гораздо чаще будут обладать особи гетерогаметного пола. Поэтому заболеваниями, которые вызываются рецессивными аллелями сцепленных с полом генов, гораздо чаще болеют мужчины, а женщины часто являются носителями таких аллелей
Неполное доминирование - это особый тип взаимодействия генных аллелей, в котором более слабый рецессивный признак не может быть полностью подавлен доминантным. В соответствии с закономерностями, открытыми Г. Менделем, целиком подавляет проявление рецессивного. Исследователь изучал ярко выраженные контрастирующие признаки у растений с проявлением либо доминантных, либо рецессивных аллелей. В некоторых случаях Мендель сталкивался с невыполнением данной закономерности, но не дал ей объяснения.
Новая форма наследования
Иногда в результате скрещивания потомки наследовали промежуточные признаки, которые не давал в гомозиготной форме родительский ген. Неполного доминирования не было в понятийном аппарате генетики до начала XX века, когда были переоткрыты законы Менделя. Тогда же многие естествоиспытатели проводили генетические эксперименты с растительными и животными объектами (томатами, бобовыми, хомяками, мышами, дрозофилами).
После цитологического подтверждения в 1902 г. Уолтером Сеттоном Менделевских закономерностей принципы передачи и взаимодействия признаков стали объяснять с позиции поведения хромосом в клетке.
В том же 1902 году Чермак Корренс описал случай, когда после скрещивания растений с белыми и красными венчиками у потомков были цветки розовой окраски - неполное доминирование. Это проявление у гибридов (генотип Аа ) признака, который является промежуточным по отношению к гомозиготному доминантному (АА ) и рецессивному (аа ) фенотипам. Подобный эффект описан для многих видов цветковых растений: львиного зева, гиацинта, ночной красавицы, ягод земляники.
Неполное доминирование — это причина изменения работы ферментов?
Механизм появления третьего варианта признака можно объяснить с позиции активности ферментов, которые по природе являются белками, а гены определяют структуру белка. У растения с гомозиготным доминантным генотипом (АА ) ферментов будет достаточно, и количество пигмента будет соответствовать норме, чтобы интенсивно окрасить клеточный сок.
У гомозигот с рецессивными аллелями гена (аа ) синтез пигмента нарушен, венчик остается неокрашенным. В случае промежуточного гетерозиготного генотипа (Аа ), доминантный ген все еще дает некоторое количество фермента, отвечающего за пигментацию, но его недостаточно для яркой насыщенной окраски. Получается цвет «наполовину».
Признаки, наследуемые по промежуточному типу
Такое неполное наследование хорошо отслеживается на признаках с вариабельным проявлением:
- Интенсивность окраски. У. Бэтсон, скрестив черных и белых кур андалузской породы, получил потомство с серебристым оперением. В определении цвета радужной оболочки глаз человека также присутствует данный механизм.
- Степень выраженности признака. Структура волос человека также определяется неполным наследованием признака. Генотип АА дает курчавые волосы, аа - прямые, а люди с обеими аллелями имеют волнистую шевелюру.
- Измеримые показатели. Длина колоса пшеницы наследуется по принципу неполного доминирования.

В поколении F2 число фенотипов совпадает с числом генотипов, что и характеризует неполное доминирование. Анализирующее скрещивание для определения гибридов не требуется, так как они внешне отличаются от доминантной чистой линии.
Расщепление признаков при скрещивании
Полное и неполное доминирование как генное взаимодействие происходит в соответствии с арифметикой законов Г. Менделя. В первом случае соотношение в F2 фенотипов (3:1) не совпадает с соотношением генотипов потомков (1:2:1), поскольку фенотипически сочетания аллелей АА и Аа проявляются одинаково. Тогда неполное доминирование - это совпадение в F2 пропорции отличающихся генотипов и фенотипов (1:2:1).
У земляники окраска я год наследуется по принципу неполного доминирования. Если скрестить растение с красными ягодами (АА ) и растение с белыми ягодами - генотип аа , то в первом поколении все полученные растения дадут плоды с розовой окраской (Аа ).

Скрестив гибриды из F1, во втором поколении F2 получим соотношение потомков, совпадающее с таковым генотипов: 1АА+2Аа+1аа . По 25% растений из второго поколения дадут красные и неокрашенные плоды, 50% растений - розовые.
Аналогичную картину будем наблюдать в двух поколениях при скрещивании чистых линий цветков ночной красавицы с венчиками пурпурного и белого цветов.

Особенности наследования в случае летальности генов
В некоторых случаях по соотношению фенотипов потомства сложно установить, как взаимодействуют гены. Во втором поколении расщепление при неполном доминировании отличается от ожидаемого 1:2:1, и от 3:1 — при полном. Это бывает, когда доминантный или рецессивный признак дает в гомозиготном состоянии фенотип, не совместимый с жизнью (летальные гены).
У каракульских овец серой окраски новорожденные ягнята, гомозиготные по доминантному аллелю окраски, погибают из-за того, что такой генотип вызывает нарушения развития желудка.
У человека пример летальности доминантной формы гена - брахидактилия (короткопалость). Признак выявляется у в случае гетерозиготного генотипа, в то время как доминантные гомозиготы погибают на ранних этапах внутриутробного развития.

Летальными могут быть и рецессивные аллели генов. Серповидно-клеточная анемия приводит, в случае появления двух рецессивных аллелей в генотипе, к изменению формы эритроцитов. Кровяные клетки не могут эффективно присоединять кислород, и 95% детей с данной аномалией погибают от кислородного голодания. У гетерозигот же измененная форма эритроцитов не влияет в такой степени на жизнеспособность.
Расщепление признаков при наличии летальных генов
В первом поколении при скрещивании АА х аа летальность не проявится, поскольку все потомки будут иметь генотип Аа. Приведем примеры расщепления признаков во втором поколении для случаев с летальными генами:
Важно понимать, что действуют при неполном доминировании обе алели, а эффект частичного подавления признака является результатом взаимодействия генных продуктов.
Алле́ли (от греч. ἀλλήλων - друг друга, взаимно) - различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологичных хромосом и определяющие альтернативные варианты развития одного и того же признака. В диплоидном организме может быть два одинаковых аллеля одного гена, в этом случае организм называется гомозиготным, или два разных, что приводит к гетерозиготному организму. Термин «аллель» предложен В. Иогансеном(1909 г.) .
Нормальные диплоидные соматические клетки содержат два аллеля одного гена (по числу гомологичных хромосом), а гаплоидные гаметы - лишь по одному аллелю каждого гена. Для признаков, подчиняющихся законам Менделя, можно рассматривать доминантные и рецессивные аллели. Если генотип особи содержит два разных аллеля (особь - гетерозигота), проявление признака зависит только от одного из них - доминантного. Рецессивный же аллель влияет нафенотип, только если находится в обеих хромосомах (особь - гомозигота). Таким образом, доминантный аллель подавляет рецессивный. В более сложных случаях наблюдаются другие типы аллельных взаимодействий (см. ниже) .
Следует, однако, отметить, что, несмотря на разнообразие взаимодействия аллелей, порой весьма сложных, все они подчиняются первому закону Менделя - закону единообразия гибридов первого поколения .
Обозначение аллелей[править | править вики-текст]
Обычно в качестве обозначения аллеля применяют сокращение названия соответствующего гена до одной или нескольких букв. В случае наличия только 2 аллелей, чтобы отличить доминантный аллель от рецессивного, первую букву в обозначении доминантного пишут заглавной.
Если же имеет место множественный аллелизм, то используется следующая система обозначения аллелей, например, в случае вышеописанной мутации ade у дрожжей. Ответственный за неё ген обозначают заглавными латинскими буквами: ADE 2 . Множественные аллели этого гена записывают так: ade 2-1 , ade 2-2 и т.д. Сам же признак, как это принято в генетике микроорганизмов, обозначают ADE .
При этом само слово «аллель» употребляется в русском языке как в женском, так и в мужском роде .
Типы аллельных взаимодействий[править | править вики-текст]
Вскоре после переоткрытия законов Менделя было установлено, что взаимодействие генов не исчерпывается одним лишь полным доминированием доминантной аллели над рецессивной. В действительности между аллелями одного гена и различными генами в генотипе существуют сложные и разнообразные взаимодействия, играющие немаловажную роль в реализацииинформации, заключённой в каждом отдельном гене .
Доминантность[править | править вики-текст]
Основная статья: Доминантность
примере цветкарододендрона (Rhododendron ).
Доминирование (доминантность) заключается в том, что один из аллелей пары (доминантный) маскирует или полностью подавляет проявление второго аллеля (рецессивного). При этом степень подавления рецессивного признака обусловливает различные варианты доминирования:
1. Полное доминирование - взаимодействие двух аллелей одного гена, когда доминантный аллель полностью исключает проявление действия второго аллеля. В фенотипе присутствует только признак, задаваемый доминантной аллелью. Например, в экспериментах Менделя пурпурная окраска цветкаполностью доминировала над белой .
2. Неполное доминирование - доминантный аллель в гетерозиготном состоянии не полностью подавляет действие рецессивного аллеля. Гетерозиготы имеют промежуточный характер признака. Например, если в гомозиготном состоянии один аллель определяет красную окраску цветка, а другой - белую, то гетерозиготный гибрид будет иметь розовые цветки . В некоторых источниках неполное доминирование характеризуют как такой тип взаимодействия аллелей, когда признак у гибридов F 1 занимает не среднее положение, а отклоняется в сторону родителя с доминирующим признаком. Полностью же средний вариант (как, например, приведённый выше пример наследования окраски цветков) относят кпромежуточному характеру наследования , то есть отсутствию доминирования .
3. Сверхдоминирование - более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной. На этом типе аллельного взаимодействия основано явление гетерозиса(превосходство над родителями по жизнеспособности, энергии роста, плодовитости, продуктивности) .
4. Кодоминирование - проявление у гибридов нового варианта признака, обусловленного взаимодействием двух разных аллелей одного гена. При этом, в отличие от неполного доминирования, оба аллеля проявляются в полной мере. Наиболее известным примером является наследование групп крови у человека . Некоторые источники также понимают именно под кодоминированием отсутствие доминантно-рецессивных отношений .
5. Доминирование, связанное с полом происходит тогда, когда одна и та же аллель у самцовпроявляется как доминантная, а у самок - как рецессивная. Например, у овцематок доминирует комолость (R), а у баранов - рогатость (R 1) .
Генные мутации.
Генные мутации
Генные (точковые) мутации затрагивают, как правило, один или несколько нуклеотидов, при этом один нуклеотид может превратиться в другой, может выпасть(делеция), продублироваться, а группа нуклеотидов может развернутся на 180 градусов. Например, широко известен ген человека, ответственный за серповидно – клеточную анемию, который может привести к летальному исходу. Соответствующий нормальный ген кодирует одну из полипептидныз цепей гемоглобина. У мутантного гена нарушен всего один нуклеотид (ГАА на ГУА). В результате в цепи гемоглобина одна аминокислота заменена на другую(вместо глутамина – валин). Казалось бы ничтожное изменение, но оно влечет за собой роковые последствия: эритроцит деформируется, приобретая серповидно – клеточную форму, и уже не способен транспортировать кислород, что и приводит к гибели организма. Генные мутации приводят к изменению аминокислотной последовательности белка. Наиболее вероятное мутация генов происходит при спаривание тесно связанных организмов, которые унаследовали мутантный ген у общего предка. По этой причине вероятность возникновения мутации повышается у детей, чьи родители являются родственниками. Генные мутации приводят к таким заболеваниям, как амавротическая идиотия, альбинизм, дальтонизм и др.
Интересно, что значимость нуклеотидных мутаций внутри кодона неравнозначна: замена первого и второго нуклеотида всегда приводит к изменению аминокислоты, третий же обычно не приводит к замене белка. К примеру, "Молчащая мутация"- изменение нуклеотидной последовательности, которая приводит к образованию схожего кодона, в результате аминокилотная последовательность белка не меняется.
Хромосомные мутации
Хромосомные мутации приводят к изменению числа, размеров и организации хромосом, поэтому их иногда называют хромосомными перестройками. Хромосомные перестройки делятся на внутри- и межхромосомные. К внутрехромосмным относятся:
· Дубликация – один из участков хромосомы представлен более одного раза.
· Делеция – утрачивается внутренний участок хромосомы.
· Инверсия –повороты участка хромосомы на 180 градусов.
Межхромосомные перестройки (их еще называют транслокации) делятся на:
· Реципрокные – обмен участками негомологичных хромосом.
· Нереципрокные – изменение положения участка хромосомы.
· Дицентрические – слияние фрагментов негомологичных хромосом.
· Центрические – слияние центромер негомологичных хромосом.
Хромосомные мутации проявляются у 1% новорожденных. Однако интересно, исследования показали, что нестабильность соматических клеток здоровых доноров не исключение, а норма. В связи с этим была высказана гипотеза о том, что нестабильность соматических клеток следует рассматривать не только как патологическое состояние, но и как адаптивную реакцию организма на измененные условия внутренней среды. Хромосомные мутации могут обладать фенотипическими явлениями. Наиболее распостраненный пример - синдром "Кошачьего крика" (плачь ребенка напоминает мяукание кошки). Обычно носители такой делеции погибают в младенчестве. Хромосомные мутации часто приводят к паталогическим нарушениям в организме, но в то же время хромосомные перестройки сыграли одну из ведущих ролей в эволюции. Так, у человека 23 пары хромосом, а у обезьяны - 24. Таким образом различие составляет всего одна хромосома. Ученые предполагают, что в процессе эволюции произошла хотя бы одна перестройка. Подтверждением этого может служить и тот факт, что 17 хромосома человека отличается от такой же хромосомы шимпанзе лишь одной перецентрической инверсией. Такие рассуждения во многом подтверждают теорию Дарвина.
Геномные мутации
Главная отличительная черта геномных мутаций связана с нарушением числа хромосом в кариотипе. Эти мутации так же подразделяются на два вида: полиплоидные анеуплоидные.
Полиплоидные мутации ведут к изменению хромосом в кариотипе, которое кратно гаплоидному набору хромосом. Этот синдром впервые был лишь обнаружен в 60-ых годах. Вообще полиплодия характерна в основном для человека, а среди животных встречается крайне редко. При полиплоидии число хромосом в клетке насчитывается по 69 (триплодие) , а иногда и по 92 (тетраплодие) хромосомы. Такое изменение ведет практически к 100 % смерти зародыша. Триплодие имеет не только многочисленные пороки, но и приводит к потере жизнеспособности. Тетраплодие встречается еще реже, но так же зачастую приводит к летальному исходу.
Анеуплоидные же мутации приводят к изменению числа хромосом в кариотипе, некратное гаплоидному набору. В результате такой мутации возникают осыби с аномальным чилом хромосом. Как и триплодия, анеуплодия часто приводит к смерти еще на ранних этапах развития зародыша. Причиной же таких последствий является утрата целой группы сцепления генов в кариотипе.
В цело же, механизм возникновения геномных мутаций связан с патологией нарушения нормального расхождения хромосом в мейозе, в результате чего образуются аномальные гаметы, что и ведет к мутации. Изменения в организме связаны с присутствием генетически разнородных клеток. Такой процесс называется мозаицизм.
Геномные мутации одни из самых страшных. Они ведут к таким заболеваниям, как синдром Дауна (трисомия, возникает с частотой 1 больной на 600 новорожденных), синдром Клайнфельтера и др.
Классификация мутаций по уровню организации наследственной информации:
- Генные (точковые ) – невидны в световой микроскоп. Причиной может быть выпадение нуклеотида, повтор одного и того же нуклеотида, замена одного нуклеотида на другой. Это всё приводит к изменению наследования нуклеотидов в молекулах ДНК. А это в свою очередь приводит к изменению последовательности аминокислот в молекуле белка.
- Хромосомные(аберрации ) – изменение макроструктуры хромосом. видны в световой микроскоп. Различают несколько видов хромосомных мутаций.
Межхромосомные (транслокации) – перенос целой хромосомы или её части и присоединение этого участка к другой негомологичной хромосоме.
Внутрихромосомные (инверсии) – выпадение участка хромосомы. Разворот этого участка хромосомы на 180 градусов и вновь встраивание в эту же хромосому.
Делеция (выпадение) – потеря срединного участка хромосомы.
Дефишенси - потеря концевого участка хромосомы.
Дупликация – повтор одного и того же участка хромосомы.
- Геномные – обусловлены изменением числа хромосом. Сами хромосомы при этом не изменяются.
Полиплоидия – увеличение числа хромосом на набор кратный гаплоидному (3n, 4n, 5n).
< Аутоплоидия – организмы, в которых умножаются хромосомы одного генома.
< Аллоплоидия – организмы, в которых умножаются хромосомы разных геномов.
Гетероплоидия – происходит изменение числа хромосом на набор не кратный гаплоидному (2n+1 - трисомия, 2n-1 - моносомия, 2n-2 - нулисомия).
Гаплоидия – в соматических клетках имеется только гаплоидный набор хромосом.
Цитоплазматические – возникают в ДНК органелл цитоплазмы.
Классификация мутаций по типу клеток, в которых возникают мутации (по локализации):
- Генеративные – возникают половых клетках и наследуются при половом размножении.
- Соматические – возникают в соматических клетках, не наследуются, но могут наследоваться у растений при вегетативном размножении.
Классификация мутаций по причине их вызывающей:
- Спонтанные – причина неизвестна.
- Индуцированные – причина известна.
Мутагены – это факторы, вызывающие мутации.